New genetic programs in Darwinism and strong panspermia
by Tom Ray / Department of Zoology / University of Oklahoma and
Brig Klyce / Astrobiology Research Trust / Memphis, Tennessee
UpdatesThe history of life on Earth is characterized by the appearance of species with new organs, parts, tissues, systems and capabilities. Examples of these new features include the first appearance of photosynthesis, oxygen metabolism, multicellularity, cell specialization, sexual reproduction, locomotion, digestive systems, circulatory systems, nervous systems, hard shells, lungs, limbs, bones, scales, feathers, skin, hair, wings, eyes, ears, etc. Such new features are made possible by new genetic programs. The genetic programs for photosynthesis and oxygen metabolism, for example, are well-studied. We would like to know where new genetic programs come from.
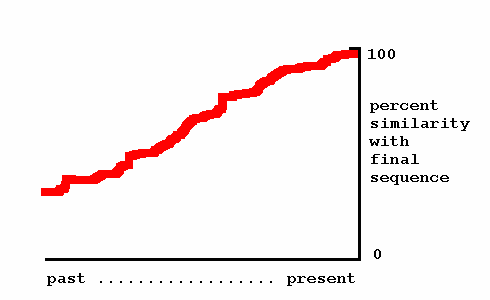
Standard Darwinian theory holds that new genetic programs arise from existing ones through gene duplication and divergence. Thus, a new program would acquire its final sequence gradually over time, as illustrated next.
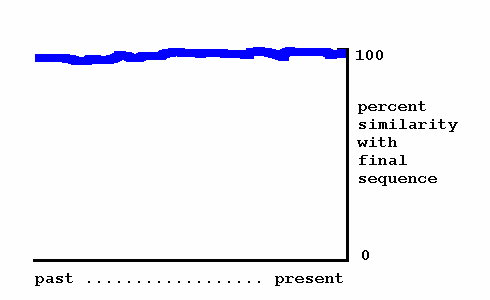
Strong panspermia begins with scepticism about this aspect of Darwinism. It is implausible to many, and direct evidence for it is thin. Meanwhile, the newly recognized possibility that microbial life may arrive from space and the growing importance of gene transfer introduce an alternative source for new genetic programs. They could be imported to Earth's biosphere and installed by gene transfer. If so, an earlier version of a genetic program would differ only slightly, if at all, from its final sequence. The progress of a genetic program over time, according to strong panspermia, is illustrated next.
Human Genome Search at University of Oklahoma
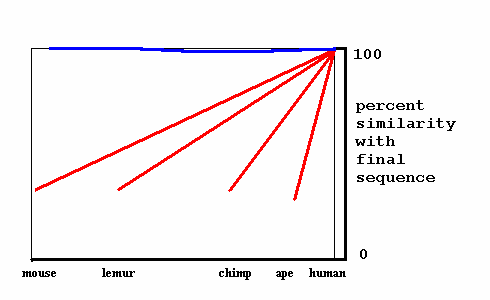
The current study by Tom Ray will begin with a comparison of the human and mouse genomes, to identify genetic programs present in humans, but absent in mice ("new genetic programs"). Then these genetic programs will be systematically sought in every available genome for species ancestral to humans that evolved after the mouse. Among these, lemur, chimp and ape genomes are substantially complete or soon will be. These intervening genomes should contain evidence of the genetic history that will provide the first picture of the process by which new genetic programs arise.
Darwinism — If a new genetic program evolves by Darwinian duplication and divergence, we should find intermediate versions of it in the intervening species between humans and mice (datapoints along one of the red lines).
Strong panspermia — If a new genetic program arrives by the strong panspermia process, intervening species should possess either nearly identical versions of it (datapoints on the blue line), or nothing similar (the program seems to have come "from nowhere".)
In spite of the limitations on the availability of genetic data, we believe that enough is available to begin work now. We anticipate that new data will be generated quickly enough to maintain the momentum of this project until a useful result is obtained.
Updates
postings by Klyce (The term "strong panspermia" has been dropped in favor of "cosmic ancestry").
 06 Nov 2025: Ancient viral DNA sequences control human embryonic development. 06 Nov 2025: Ancient viral DNA sequences control human embryonic development.
11 Apr 2025: ...complete, phased, diploid genomes of six ape species....
29 Nov 2023: ...hundreds of thousands of regulatory elements that emerged very recently
22 Mar 2023: ...de novo genes that influence the growth of the human brain.
 "De novo genes with an lncRNA origin encode unique human brain developmental functionality," by N.A. An, J. Zhang, F. Mo et al., doi:10.1038/s41559-022-01925-6, Nat Ecol Evol, 02 Jan 2023. "De novo genes with an lncRNA origin encode unique human brain developmental functionality," by N.A. An, J. Zhang, F. Mo et al., doi:10.1038/s41559-022-01925-6, Nat Ecol Evol, 02 Jan 2023.
"Evolution: Miniproteins appeared 'from nowhere'," The Max Delbrück Center for Molecular Medicine [link | alt.], 17 Feb 2023.
"Novel integrative elements and genomic plasticity in ocean ecosystems," by T. Hackl et al., doi:10.1016/j.cell.2022.12.006, Cell, 05 Jan 2023.
Molecular and cellular evolution of the primate dorsolateral prefrontal cortex, by Shaojie Ma, Mario Skarica et al., doi:10.1126/science.abo7257, Science, 25 Aug 2022.
16 May 2022: The gene regulatory network controlling the development of germ cells may have been rewired by ERV insertions during hominoid evolution.
Evolutionary impact of chimeric RNAs on generating phenotypic plasticity in human cells, by Sumit Mukherjee and Milana Frenkel-Morgenstern, doi:10.1016/j.tig.2021.08.015, Trends in Genetics, 01 Jan 2022.
"A high-quality bonobo genome refines the analysis of hominid evolution", by Yafei Mao et al., doi:10.1038/s41586-021-03519-x, Nature, 05 May 2021. We classify 15,786 insertions and 7,082 deletions [>50 bp] as bonobo-specific....
"Synteny-based analyses indicate that sequence divergence is not the main source of orphan genes" [link], by Nikolaos Vakirlis, Anne-Ruxandra Carvunis and Aoife McLysaght, doi:10.7554/eLife.53500, eLife, 18 Feb 2021.
Evolution of the human brain by Colette Dehay and Henry Kennedy, doi:10.1126/science.abd1840, Science, 31 Jul 2020. A human-specific gene is a determinant of the cognitive architecture of the human cerebral cortex.
27 May 2019: de novo gene birth
21 Sep 2018: De novo genes continue to confound standard darwinism.
01 Jun 2018: New genes increase the size of the human brain.
15 May 2018: ...many genes typically associated with metazoan functions actually pre-date animals themselves....
|
What about the big bang? We are aware of the assertion that genetic programs must originate at some time. The foundation for this claim is the big bang. However, if new genetic programs do originate as Darwinism prescribes, the process would have continued at least until the emergence of the human species. It should be possible to obtain evidence of this recent process. If the evidence is obtained, the existing paradigm would get welcome confirmation. If direct evidence is not obtained after sustained effort, that result would also contribute to science.
The situation was analogous in the 1880s, when Al Michelson sought direct evidence of the luminiferous aether. Almost no one doubted its existence. The highly successful wave theory of light seemed to require it. The definitive experiment was performed by Michelson and Ed Morely, and reported in 1887. When no evidence of the aether was found, a major scientific advance ensued. |
Transposable elements are the primary source of novelty in primate gene regulation by Marco Trizzino et al., Genome Res., online 30 Aug 2017.
A Surprise Source of Life's Code by Emily Singer, Quanta Magazine (+Scientific American), 18 Aug 2015. Of the 600 human-specific genes that [evolutionary biologist Mar] Albà's team found, 80 percent are entirely new, having never been identified before.
27 Sep 2017: ...multiple waves of LINE retrotransposition and the birth of new mobile elements during primate evolution, and their impact on the evolving human brain.
Using the Neandertal genome to study the evolution of small insertions and deletions in modern humans by Manjusha Chintalapati et al., BMC Evolutionary Biology, 04 Aug 2017.
09 Apr 2016: The human reference genome contains over 120 HML-2 [endogenous retrovirus] insertions that are not present in chimpanzees, bonobos, or gorillas.
10 Mar 2016: Viruses... have had a significant impact on the evolution of all organisms, from bacteria to humans.
03 Mar 2016: ...These viral fossils probably played a key role in the evolution of our species....
10 Dec 2015: ...The arrival of this new group [diplobasts] was accompanied by a small number of novel proteins (specified by previously nonexistent genes)....
4 Sep 2015: ...Thousands of transcripts ...likely to have originated de novo...., and
4 Jan 2016: Update about de novo transcripts.
Marta Florio et al., "Human-specific gene ARHGAP11B promotes basal progenitor amplification and neocortex expansion " [abstract], doi:10.1126/science.aaa1975, p 1465-1470 v 347, Science, 27 Mar 2015. "...56 genes that lack mouse orthologs.... 207 human genes with mouse orthologs...."
7 Mar 2015: ...For most of the functional signature systems of the eukaryotic cells, we can detect no evolutionary intermediates. — Eugene V. Koonin
13 Jan 2015: ...Inherited viruses that are millions of years old play an important role in building up the complex networks that characterise the human brain.
8 Oct 2014: 24 hominoid-specific de novo protein-coding genes were identified. (Points along the blue line)
10 Sep 2014: Reconstruction of evolution ...demonstrates that extensive gene loss and horizontal gene transfer leading to innovation are the two dominant evolutionary processes....
8 Jun 2013: Where is the evidence for new genetic programming by standard darwinian trial-and-error?
11 May 2013: ...TEs, and in particular ERVs, have contributed hundreds of thousands of novel regulatory elements to the primate lineage....
Macarena Toll-Riera and M Mar Albà, "Emergence of novel domains in proteins" [abstract | pdf], doi:10.1186/1471-2148-13-47, n47 v13, BMC Evolutionary Biology, 20 Feb 2013. "We conclude that proteins tend to gain domains over time....."
25 Jan 2013: Many of our genes have no obvious relatives or evolutionary history. So where did they come from?
23 Nov 2012: Some regulatory sequences in the human genome have no identifiable orthologs in chimpanzee, gorilla, orangutan, rhesus macaque, marmoset, or any of six other species investigated.
Wesley C. Warren et al., "Genome analysis of the platypus reveals unique signatures of evolution" [abstract], doi:10.1038/nature06936, p175-183 v453, Nature, 8 May 2008. "As expected, the majority of platypus genes (82%; 15,312 out of 18,596) have orthologues in these five other amniotes. The remaining 'orphan' genes are expected to primarily reflect rapidly evolving genes, for which no other homologues are discernible, erroneous predictions, and true lineage-specific genes that have been lost in each of the other five species under consideration."
23 Oct 2012: Evolution by subfunctionalization
Shengjun Tan et al., "Variation of presence/absence genes among Arabidopsis populations" [Open Access abstract], doi:10.1186/1471-2148-12-86, v12 n86, BMC Evolutionary Biology, online 14 Jun 2012. "Using our approach, 2,407 genes were identified with P/A polymorphisms, indicating that about 10% of genes are likely to be maintained as P/A genes in Arabidopsis. Both the number and the proportion of P/A genes are remarkable, which may reflect their important roles in the species adaptation to the environment."
Cheryl P Andam et al., "Ancient origin of the divergent forms of leucyl-tRNA synthetases in Halobacteriales" [Open Access abstract], doi:10.1186/1471-2148-12-85, v12 n85, BMC Evolutionary Biology, online 13 Jun 2012. "The study ...illustrates the importance of gene transfer originating in lineages that went extinct since the transfer occurred." Translation: the genes seem to have come from nowhere.
9 Mar 2012: ...Fundamental elements of the C4 pathway ...were acquired via a minimum of four independent lateral gene transfers....
Alfonso Natale et al., "Evolution of anterior Hox regulatory elements among chordates" [Open Access abstract], doi:10.1186/1471-2148-11-330, v11 n330, BMC Evolutionary Biology, online 15 Nov 2011. "This analysis revealed that Kreisler and Krox20 dependent enhancers critical in segmental regulation of the hindbrain appear to be specific for the vertebrate lineage. In contrast, neural enhancers that function as Hox response elements through the action of Hox/Pbx binding motifs have been conserved during chordate evolution."
24 Oct 2011: New genes are preferentially expressed... (genes are either Young, or Old.)
26 Sep 2011: More than 1500 genes necessary for the evolution of pregnancy in placental mammals were recruited into service....
26 Aug 2011: ...Adaptive introgression of archaic alleles has significantly shaped modern human immune systems.
25 Oct 2010: Genes are either very old, or they appear suddenly, without predecessors.
20 Sep 2010: Eukaryotic genes come from archaebacteria or eubacteria... or not.
13 Sep 2010: Origins, evolution, and phenotypic impact of new genes, a recent review deconstructed following the above criteria.
 The Independent Evolution of Multicellularity, a new webpage, notes that the test proposed above is universally applicable, 23 Jun 2010. The Independent Evolution of Multicellularity, a new webpage, notes that the test proposed above is universally applicable, 23 Jun 2010.
17 Feb 2010: We suggest that both ...were derived from ancestral bacterial proteins....
21 Sep 2009: Nearly half of the human genome is derived from transposable elements (TEs).
17 Sep 2009: The gain and loss of exons has contributed to the evolution of new features.
Three New Human Genes is a related new CA webpage, posted 4 Sep 2009.
23 Jul 2009: Primate-specific genes were inserted de novo, not generated by gradual divergence from non-primate genes.
30 Oct 2008: Horizontal transfer (HT) of DNA transposons has apparently produced evolutionary innovation in mammals.
22 Aug 2008: It is now completely clear that genomic complexity was present very early on....
21 Apr 2008: Placental genes have ancient origins.
29 Nov 2007: Surprises come from a comparison of twelve fruitfly genomes.
19 Nov 2007: Ancient retroviruses spurred evolution of gene regulatory networks in humans and other primates.
26 Sep 2007: The genomes of 17 species of fungi have been analysed to reconstruct gene duplication....
26 Aug 2007: The first analysis of the genome of the sea anemone shows it to be nearly as complex as the human genome.
Xun Gu, "Evolutionary Framework for Protein Sequence Evolution and Gene Pleiotropy" [abstract], doi:10.1534/genetics.106.066530, p 1813-1822 v 175, Genetics, Apr (online 4 Feb) 2007.
15 Sep 2006: Tom Ray replies to our open email of 31 Dec 2005. (The project went nowhere.)
4 May 2006: A gene captured from a mobile element fused with another gene to make a new primate gene.
Zhixi Su et al., "Evolution of alternative splicing after gene duplication" [abstract], doi:10.1101/gr.4197006, Genome Research, online 19 Dec 2004. "...Loss of alternative splicing in duplicate genes may occur shortly after the gene duplication. These results support the subfunctionization model of alternative splicing in the early stage after gene duplication."
30 Sep 2005: The chimp genome has been sequenced. At least seventeen human genes contain exons missing in chimps.
27 June 2005: "Gene duplication is the primary source of new genes."
21 Nov 2004: Evidence that 1,183 human genes were "born" 3-4 million years ago, by duplication and divergence....
21 Oct 2004: Eukaryote-to-eukaryote lateral gene transfer....
Elizabeth Pennisi, "Surveys Reveal Vast Numbers of Genes" [summary], p 1591 v 304, Science, 11 June 2004.
Correspondence with Tom Ray... during the establishment and pursuit of this research project, 2001-2005.
7 May 2004: Ultraconserved elements.
15 Jan 2004: Are normal microevolutionary processes sufficient to account for human origins?
2003, November 20: In mammals, CNGs are more numerous and better conserved than genes — some datapoints on the blue line!
Protein Evolution.doc: (943KB) interim report from Thomas Ray, 29 July 2003.
2003, February 18: Humanoid gene arose abruptly?
2002, November 24: Photosynthesis evolved by gene transfer.
2001 Nov 18: Human Genome Search at U of Oklahoma, the study by Tom Ray.
|